The predominate working hypothesis in the
psychophysical literature has been a centralized internal clock
model (Creelman 1962, Treisman 1963; for a review see Allan
1979), in which an oscillator beating at a fixed frequency
generates tics that are detected by a counter. These models
often assume that timing is centralized, that is, the brain
uses the same circuitry to determine the duration of an
auditory tone and for the duration of a visual
flash.
The basic computational unit of clock theories
involves an oscillator and a counter (Creelman 1962,
Treisman 1962). Conceptually, the oscillator beats at some
constant frequency, and each beat would then be counted by some
sort of neural integrator. These ideas have not yet been
expressed concretely in terms of the synaptic organization of a
specific brain region.
However, as proposed by Meck and colleagues,
clock-like mechanisms could be involved in timing on the scale of
seconds and minutes (Meck 1996, Matell & Meck 2000).
.....
the dominant model of temporal processing is the internal clock
model. A prototypical clock model includes an oscillator
(pacemaker) that emits pulses that are counted by an
accumulator (Creelman, 1962;
Treisman, 1963; Church, 1984; Gibbon et al., 1997).
Within this framework, the pulse count provides a linear metric
of time, and temporal judgments rely on comparing the current pulse
count to that of a reference time. This model has proven
effective in providing a framework for much of the psychophysical
data relating to temporal processing (Church, 1984; Meck, 1996; Rammsayer and Ulrich,
2001). However, electrophysiological and
anatomical support for the putative accumulator remains
elusive, and mounting evidence indicates that clock models are
not entirely consistent with the experimental data (for reviews see
Mauk and Buonomano, 2004; Buhusi and
Meck, 2005).
The standard model of temporal processing
postulates a single centralized internal clock, which relies on an
oscillator and an accumulator (counter) (Creelman, 1962; Treisman, 1963; Church, 1984;
Grondin, 2001). The clock concept is generally
taken to imply that the passage of time is counted in units that
can be combined or compared linearly.
Traditionally, the manner in which
durations in the seconds-to-minutes range are perceived,
represented and estimated has been explained using a
pacemaker–accumulator model. This model is relatively
straightforward, and provides powerful explanations of both
behavioural and physiological data. However, recent advances
that challenge the traditional pacemaker–accumulator
model have come from studies that use various modern
techniques, which range from drug microinjection and ensemble
recording in genetically modified and wild- type rodents to
functional MRI (fMRI) and positron emission tomography (PET) in
neurologically impaired and control humans. These data indicate
that time might be represented in a distributed manner in the
brain, and that telling the time is a matter of detecting the
coincidental activation of different neural populations.
Human behavior based on the perception and timing
in the range of seconds-to- minutes has traditionally been
explained bythe pacemaker–accumulator model.
The
pacemaker- accumulator model (PAM), which is based on
scalar
expectancy or timing theory (Church, 1984; Gibbon
et al., 1984; Meck, 1983), “is relatively straightforward,
and provides powerful explanations of both behavioural and
physiological data” (Buhusi & Meck, 2005; p.
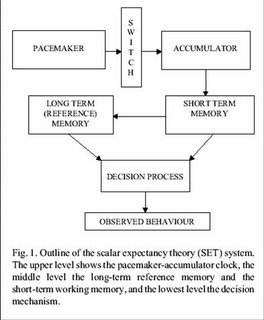
755).Briefly, the PAM model implicates the processing
of temporal information via three synchronized modular
information processing systems (see Buhusi & Mech, 2005.)
The “clock” system consists of a dopaminergic
pacemaker that regularly generates or emits neural ticks or pulses
that are transferred (via a “gaiting” switch) to the
accumulator, which accumulates ticks/pulses (neural counting) that
correspond to a specific time interval. The raw representation
of the stimulus duration in the accumulator is then transferred to
working memory, a component of the PAM “memory”
system. The contents of working memory are then compared against a
“reference standard” in the long-term
(reference) memory, the second component of the PAM memory system.
Finally, the “decision” level of the PAM is
conceptualized to consist of a comparator that determines an
appropriate response based on a decision rule which involves a
comparison between the interval duration value present in working
memory and the corresponding duration value in reference memory. In
other words, a comparison is made between the contents of reference
memory (the standard) and working memory (viz., are they
“close?”).
Various models of time perception have been
suggested, the most popular being the internal- clock model
(Gibbon, 1977). Here a series of pulses are produced by an
internal pacemaker; these pulses are collated, counted and then
compared to stored representations in order to allow the brain to
judge durations and produce time estimations. Such models have been
extensively studied using behavioural paradigms (Thomas
&Weaver, 1975; Block, 1990).
Internal clock models based on neural counting
provide a useful heuristic for explaining human performance on the
temporal discrimination of brief intervals. It is not
surprising, therefore, that the notion of a pacemaker-counter
system represents a fundamental feature of most psychophysical
models of temporal discrimination introduced over the last 4
decades (e.g., Allan, Kristofferson, & Wiens, 1971;
Creelman, 1962; Gibbon, 1977; Killeen & Fetterman, 1988;
Penton-Voak, Edwards, Percival, & Wearden, 1996; Treisman,
1963; Treisman, Faulkner, Naish, & Brogan, 1990).


